ntegrative veterinary medicine.
ntegrative veterinary medicine.
-
NAD+ COMO EIXO CENTRAL DA BIOENERGIA CELULAR: FONTES NUTRICIONAIS NATURAIS, DISPOSIÇÃO ENERGÉTICA E ESTRATÉGIAS INTEGRATIVAS NA ALIMENTAÇÃO NATURAL PARA CÃES E GATOS
INSTITUTO PETCLUBE AMICHETTI DE MEDICINA VETERINÁRIA INTEGRATIVA
NAD+ COMO EIXO CENTRAL DA BIOENERGIA CELULAR: FONTES NUTRICIONAIS NATURAIS, DISPOSIÇÃO ENERGÉTICA E ESTRATÉGIAS INTEGRATIVAS NA ALIMENTAÇÃO NATURAL PARA CÃES E GATOS
"O NAD+ não é apenas uma coenzima — é a ponte entre o alimento que o animal consome e a energia que suas células utilizam."
16 de maio de 2026
Autores: Dr. Cláudio Amichetti Júnior – Médico Veterinário Integrativo (CRMV-SP 75.404 VT, MAPA 00129461/2025), Foco em Nutrição Clínica, Medicina Canabinoide e Alimentação Natural. Dr. Gabriel Amichetti – Médico Veterinário (CRMV-SP 45.592 VT), Especialista em Ortopedia e Cirurgia de Pequenos Animais.
RESUMO
"Sem NAD+ suficiente, a célula repara pior, regula pior, protege pior e produz energia de forma ruim."
O Nicotinamida Adenina Dinucleotídeo (NAD+) é uma coenzima central da bioenergia celular, atuando como mediador essencial no metabolismo energético e na ativação de sirtuínas. Este artigo revisa o papel do NAD+ na disposição energética de cães e gatos, abordando seu declínio fisiológico associado ao envelhecimento e ao estresse metabólico. São discutidas as "três grandes máquinas" consumidoras de NAD+: sirtuínas (longevidade), PARPs (reparo de DNA) e a enzima CD38 (degradação inflamatória). A análise destaca a ineficácia da suplementação oral direta e a superioridade da alimentação natural como fonte de precursores biodisponíveis, como niacina, triptofano e nucleotídeos. Em contrapartida, a ração comercial extrusada apresenta defasagem nutricional devido à degradação térmica de vitaminas do complexo B. Por fim, explora-se a sinergia do NAD+ com peptídeos mitocondriais (SS31 e MOTS-c) no reparo da função mitocondrial, oferecendo uma abordagem integrativa para a promoção da saúde e longevidade na clínica de pequenos animais.
Palavras-chave: NAD+. Disfunção mitocondrial. Alimentação natural. Sirtuínas. Medicina veterinária integrativa.
ABSTRACT
"Without sufficient NAD+, the cell repairs worse, regulates worse, protects worse, and produces energy poorly."
Nicotinamide Adenine Dinucleotide (NAD+) is a central coenzyme of cellular bioenergetics, acting as an essential mediator in energy metabolism and sirtuin activation. This article reviews the role of NAD+ in the energy status of dogs and cats, addressing its physiological decline associated with aging and metabolic stress. The "three major machines" that consume NAD+ are discussed: sirtuins (longevity), PARPs (DNA repair), and the CD38 enzyme (inflammatory degradation). The analysis highlights the inefficiency of direct oral supplementation and the superiority of natural feeding as a source of bioavailable precursors such as niacin, tryptophan, and nucleotides. In contrast, commercial extruded kibble shows nutritional deficiency due to thermal degradation of B-complex vitamins. Finally, the synergy of NAD+ with mitochondrial peptides (SS31 and MOTS-c) in repairing mitochondrial function is explored, offering an integrative approach to promoting health and longevity in small animal practice.
Keywords: NAD+. Mitochondrial dysfunction. Natural feeding. Sirtuins. Integrative veterinary medicine.
1 INTRODUÇÃO
"O NAD+ é a molécula central da bioenergia — sem ela, a usina mitocondrial para, e o animal paga o preço com fadiga, inflamação e envelhecimento precoce."
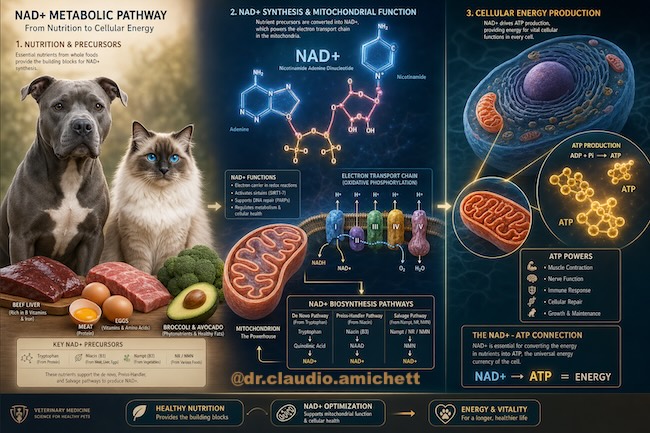
O NAD+ (Nicotinamida Adenina Dinucleotídeo) é uma coenzima essencial presente em todas as células vivas. Conforme destaca Cardoso (2025), "o NAD+ não é um peptídeo, é uma coenzima, uma molécula central da bioenergia" [1]. Sua função primária é atuar como aceptor de elétrons em reações de oxirredução, permitindo a conversão de macronutrientes em energia química utilizável na forma de Adenosina Trifosfato (ATP). Sem a presença onipresente desta molécula, a vida aeróbica como a conhecemos seria impossível, uma vez que ela sustenta o fluxo eletrônico necessário para a homeostase celular.
Descoberto por Harden e Young em 1906, o NAD+ foi posteriormente elucidado por Otto Warburg como peça-chave na respiração celular [2]. Ele funciona como um "cabo de transmissão" que conecta o ciclo de Krebs à fosforilação oxidativa, viabilizando a produção massiva de ATP [3]. Na medicina veterinária contemporânea, a compreensão desta coenzima transcende a bioquímica básica, tornando-se um marcador fundamental de vitalidade e um alvo terapêutico para doenças degenerativas crônicas em pequenos animais.
A "disposição energética" de cães e gatos depende da capacidade do organismo em extrair energia dos alimentos e distribuí-la para todas as funções biológicas: contração muscular, termorregulação, função cognitiva, metabolismo hepático e resposta imune. Todas essas funções demandam ATP, e a produção de ATP é diretamente dependente da disponibilidade de NAD+ na mitocôndria. Como ressalta Cardoso (2025), "sem NAD+ suficiente no seu corpo, a célula vai reparar pior, vai regular pior, vai proteger o seu corpo pior e produzir de uma forma ruim a energia" [1]. Um cão ou gato com baixos níveis de NAD+ invariavelmente apresentará menor disposição, fadiga crônica, recuperação lenta e declínio funcional, muitas vezes mascarados sob o rótulo de senilidade inevitável.
A alimentação natural (AN) para cães e gatos, baseada em ingredientes frescos e minimamente processados, oferece vantagens significativas para a manutenção dos níveis de NAD+. Alimentos como fígado bovino, coração, sardinha, ovos e carnes frescas são ricos em niacina (vitamina B3), triptofano, riboflavina (B2) e piridoxina (B6) — precursores e cofatores essenciais para a biossíntese de NAD+. Além disso, a matriz alimentar complexa da AN favorece a absorção sinérgica desses nutrientes, respeitando a fisiologia de carnívoros facultativos e estritos [4].
A ração comercial extrusada passa por processamento em altas temperaturas (120-180°C) que degrada significativamente as vitaminas do complexo B, especialmente a tiamina (B1), riboflavina (B2) e niacina (B3). O triptofano, aminoácido essencial precursor do NAD+ pela via De Novo, também sofre redução durante a extrusão [5]. Além disso, as rações comerciais não fornecem cofatores enzimáticos como Coenzima Q10 e carnitina em quantidades significativas. Vale destacar que a suplementação oral de NAD+ puro é ineficaz — a molécula é grande demais e é degradada pelo ácido gástrico antes de chegar à corrente sanguínea [3]. A melhor estratégia é fornecer os precursores corretos através da dieta fresca.
Este artigo revisa a bioquímica do NAD+, suas vias de biossíntese, as fontes naturais na alimentação para pets, as formas de produção de energia e os mecanismos de defesa anti-inflamatória mediados por esta coenzima fundamental, oferecendo ao clínico veterinário ferramentas para abordar a disfunção mitocondrial como causa raiz do envelhecimento e das doenças crônicas.
2 BIOQUÍMICA DO NAD+: MECANISMOS E FUNÇÕES CELULARES
"O NAD+ não é um produto final — é o cabo que transmite energia bruta para dentro da mitocôndria, e sem ele a usina celular trava."
A estrutura do NAD+ consiste em dois nucleotídeos unidos por seus grupos fosfato, sendo um deles a nicotinamida e o outro a adenosina. Sua importância reside na capacidade de alternar entre o estado oxidado (NAD+) e o estado reduzido (NADH). Esta transição é o motor das reações redox celulares, expressa pela equação simplificada:
NAD++H++2e−⇌NADH
No ciclo de Krebs, o NAD+ atua captando elétrons resultantes da oxidação de carboidratos, gorduras e proteínas. O NADH gerado segue para a cadeia de transporte de elétrons (CTE) na membrana interna mitocondrial. Cardoso (2025) explica que "o NAD+ é a forma bruta necessária nas mitocôndrias para gerar ATP, enquanto o NADH entrega a energia de uma forma mais constante. Para que o NADH funcione, não pode tomar ele sozinho — precisa dos dois em conjunto" [1]. Este equilíbrio é o que define o potencial redox da célula.
Na prática clínica, quando não há NAD+ disponível para captar elétrons, todas as enzimas do ciclo de Krebs ficam travadas e a produção de energia simplesmente não acontece. O resultado é uma falência energética subclínica que se manifesta como cansaço, fadiga e névoa mental no paciente veterinário [1]. A mitocôndria, incapaz de processar o fluxo de elétrons, passa a gerar um excesso de espécies reativas de oxigênio (EROS), danificando suas próprias membranas e o DNA mitocondrial.
A segunda grande função do NAD+ é a sinalização celular através da ativação das sirtuínas. Cardoso (2025) descreve as sirtuínas como "o time da longevidade — mestres da manutenção e adaptação celular" [1]. Estas enzimas dependem exclusivamente do NAD+ para remover grupos acetil de proteínas, regulando a expressão gênica e o metabolismo. As principais isoformas são:
1. SIRT1: Localizada no núcleo, promove o reparo do DNA e ativa o PGC-1α, estimulando a biogênese mitocondrial.
2. SIRT3: Localizada na matriz mitocondrial, reduz o vazamento de elétrons e protege contra o estresse oxidativo.
3. SIRT6: Crucial para a proteção dos telômeros e a estabilidade genômica.Cardoso (2025) também descreve as "três grandes máquinas que gastam NAD+": as sirtuínas, as PARPs e a CD38 [1]. As PARPs (Poli-ADP-ribose polimerases) funcionam como "mecânicos do DNA" — quando detectam dano genômico, consomem grandes quantidades de NAD+ para realizar o reparo. Em animais expostos a dietas inflamatórias e sedentarismo, o dano ao DNA é constante, levando a um esgotamento crônico do NAD+ pelas PARPs. Já a CD38 é o "ladrão do NAD+", uma enzima que degrada a coenzima e cujos níveis aumentam dramaticamente com processos inflamatórios crônicos (elevando IL-6, TNF-α, PCR), criando um ciclo vicioso de depleção energética e inflamação [1].
3 DECLÍNIO DO NAD+ COM O ENVELHECIMENTO E IMPLICAÇÕES CLÍNICAS
"A queda do NAD+ é fisiológica — envelhecer é, em grande parte, perder a capacidade de produzir energia."
Cardoso (2025) afirma categoricamente: "a queda é fisiológica — com o envelhecimento é normal que os níveis de NAD+ caiam. Quanto mais velho o animal vai ficando, menos NAD+ ele tem. E isso piora com a inflamação, dano e estresse metabólico" [1]. Este declínio não é apenas um marcador de idade, mas um motor do processo de senescência. Em cães e gatos, esse fenômeno explica por que animais idosos perdem massa muscular (sarcopenia) e apresentam menor tolerância térmica.
Existem três mecanismos principais para este declínio: (1) a via NAMPT de salvamento perde eficiência, reduzindo a reciclagem da nicotinamida; (2) o aumento do dano ao DNA recruta excessivamente as PARPs; e (3) a inflamação sistêmica de baixo grau (inflammaging) superestimula a CD38 [1, 6]. O resultado é uma "crise energética" onde a demanda por reparo supera a capacidade de produção de NAD+.
Os tecidos com alta demanda metabólica são os primeiros a sofrer. No músculo, a falta de NAD+ leva à disfunção mitocondrial e perda de força. No fígado, contribui para a resistência à insulina e esteatose. No coração, compromete a contratilidade miocárdica. Clinicamente, observamos animais que "desistem" de caminhadas longas, apresentam tremores musculares e dificuldade em manter o peso ideal mesmo com ingestão calórica adequada [1, 7].
Cardoso (2025) conecta o NAD+ baixo ao conceito de "brain rot" (cérebro envelhecido): "a ativação de sirtuínas protege melhor o sistema nervoso. O NAD+ baixo afeta a proteção e resiliência neuronal" [1]. A deficiência está ligada ao acúmulo de proteína beta-amiloide e ao aumento de NFL (Neurofilament Light Chain), um marcador de dano neuronal. Em cães, isso se traduz na Síndrome de Disfunção Cognitiva (SDC), onde o animal perde a orientação espacial e altera o ciclo sono-vigília [1].
4 VIAS DE BIOSSÍNTESE DO NAD+ E SUAS FONTES NATURAIS
"O corpo produz, recicla e gasta NAD+ — e a alimentação natural é a ferramenta mais potente para sustentar esse equilíbrio."
4.1 Via de Preiss-Handler
Esta via utiliza o ácido nicotínico (niacina) proveniente da dieta. Através da enzima NAPRT, o ácido nicotínico é convertido em NAD+. É uma via altamente eficiente. As fontes naturais mais ricas para cães e gatos incluem o fígado bovino, coração, rim, sardinha e salmão. A inclusão de vísceras na alimentação natural garante o aporte necessário para esta via [4].
4.2 Via de Salvamento (Reciclagem)
Responsável por mais de 85% do NAD+ diário, esta via recicla a nicotinamida (NAM) gerada pelas enzimas que consomem NAD+. Cardoso (2025) explica: "a NAMPT é a enzima que ajuda a transformar o material usado em NAD+ útil de novo. Só que com a idade, essa via perde força" [1]. Esta via é potencializada pelo exercício físico moderado, que aumenta a expressão de NAMPT no tecido muscular.
4.3 Via De Novo (Triptofano)
O organismo pode sintetizar NAD+ a partir do aminoácido essencial triptofano, embora a eficiência seja baixa (1-2%). Esta via é extremamente dependente de cofatores: vitamina B6, B2 e Ferro. Fontes como ovos, peru e carnes frescas fornecem o triptofano necessário, mas a deficiência de B6 (comum em dietas ultraprocessadas) bloqueia esta conversão [9].
4.4 Precursores Diretos
O Nicotinamida Ribosídeo (NR) e o Nicotinamida Mononucleotídeo (NMN) são intermediários que entram na célula de forma mais direta. O NR é encontrado no leite cru, enquanto o NMN está presente em vegetais como abacate, brócolis e pepino [8]. Embora carnívoros consumam poucos vegetais, a inclusão de pequenas porções trituradas na AN pode oferecer suporte adicional de NMN.
5 ALIMENTAÇÃO NATURAL COMO ESTRATÉGIA NUTRICIONAL PARA SUPORTE AO NAD+
"Shape é consequência de bons hábitos — e a mitocôndria responde primeiro à qualidade do que o animal ingere."
A superioridade da alimentação natural (AN) reside na preservação da integridade dos micronutrientes. Enquanto a ração comercial passa por extrusão em altas temperaturas, degradando 30-60% das vitaminas do complexo B, a AN mantém os precursores em sua forma mais biodisponível [5]. A presença de enzimas naturais e a ausência de conservantes sintéticos reduzem a carga inflamatória sistêmica, preservando o NAD+ que seria gasto pela CD38.
As vísceras musculares e secretoras são "superalimentos" mitocondriais. O coração é uma fonte inigualável de Coenzima Q10 e niacina, enquanto o fígado fornece o espectro completo de vitaminas B. Estes nutrientes atuam sinergicamente: a niacina provê o esqueleto do NAD+, enquanto a riboflavina (B2) é precursora do FAD, outro transportador de elétrons vital. A AN permite que o clínico ajuste a dieta para fornecer densidade nutricional sem excesso calórico [9].
Cardoso (2025) enfatiza: "de nada adianta usar peptídeo ou coenzima se a base não estiver correta — dieta, treino e sono precisam estar alinhados. Shape é consequência de bons hábitos" [1]. Na veterinária, o "treino" equivale ao enriquecimento ambiental e passeios ativos, e o "sono" à higiene do descanso do pet. Sem esses pilares, a biossíntese de NAD+ via NAMPT permanece deprimida.
Para pacientes geriátricos, a suplementação oral de NAD+ puro é ineficaz devido à degradação gástrica. A estratégia de ouro é a "suplementação via matriz alimentar": oferecer alimentos frescos ricos em precursores. Em casos de disfunção severa, o uso de precursores como NMN ou NR manipulados em veículos que protejam a molécula pode ser considerado, mas sempre sobre a base de uma dieta fresca [3].
6 FORMAS DE PRODUÇÃO DE ENERGIA E DEFESA ANTI-INFLAMATÓRIA DO NAD+
"Sem NAD+ suficiente, a usina mitocondrial não para totalmente, mas perde eficiência e gera uma baderna no metabolismo."
O NAD+ é o mediador entre o catabolismo e o anabolismo. Quando os níveis estão baixos, o ciclo de Krebs perde velocidade, a CTE não recebe elétrons suficientes e o gradiente de prótons enfraquece. Cardoso (2025) explica: "NAD+ baixo piora a oxidação de gordura e aumenta a produção de radicais livres. Gera uma baderna no metabolismo" [1]. O animal entra em um estado de "fome celular" mesmo estando obeso, pois não consegue converter a gordura estocada em ATP de forma eficiente.
A defesa anti-inflamatória do NAD+ é mediada principalmente pelas sirtuínas. A SIRT1 inibe o complexo NF-κB, o principal interruptor da inflamação. A SIRT3 limpa os radicais livres dentro da mitocôndria. Quando o NAD+ está alto, a inflamação é mantida sob controle. Quando cai, a inflamação crônica se instala, ativando a CD38, que por sua vez consome ainda mais NAD+ [10].
Cardoso (2025) alerta para o perigo de viver com o corpo inflamado: "a inflamação acelera o esgotamento de NAD+" [1]. Na clínica veterinária, isso é visto em animais com alergias crônicas, otites de repetição e obesidade. A alimentação natural quebra este ciclo ao fornecer antioxidantes naturais (polifenóis e flavonoides) que reduzem a ativação da CD38, poupando o NAD+ para funções de reparo e energia.
7 SINERGIA ENTRE NAD+ E PEPTÍDEOS MITOCONDRIAIS NA PRÁTICA INTEGRATIVA
"NAD+ abastece a mitocôndria — SS31 e MOTS-c a estruturam. Juntos, atacam a causa raiz do envelhecimento."
A eficácia do NAD+ depende da integridade da membrana mitocondrial. A cardiolipina é o fosfolipídio que mantém os complexos da CTE organizados. Com o tempo, a cardiolipina oxida, desorganizando a produção de energia. Cardoso (2025) recomenda a sinergia: "NAD+ casa lindamente bem com MOTS-c e SS31 para reparo e otimização mitocondrial" [1]. O SS31 protege a cardiolipina, enquanto o MOTS-c regula o metabolismo da glicose e a resposta ao estresse.
É fundamental diferenciar NAD+ de NADH. "O NAD+ é a forma bruta para dar o tranco nas mitocôndrias. O NADH faz um equilíbrio de energia mais constante, mas só funciona se houver energia prévia. Por isso, tomar NADH sozinho muitas vezes não melhora a disposição — precisa dos dois em conjunto" [1]. Na prática integrativa, o suporte deve visar o aumento do pool total de NAD+.
O tempo de resposta biológica é de 3 a 6 meses. "São meses para mudar hábitos e ver a restauração mitocondrial" [1]. O clínico deve educar o tutor que a restauração da vitalidade do pet não é imediata, mas um processo de reconstrução celular que exige consistência na dieta e no manejo de vida.
8 CONSIDERAÇÕES FINAIS
"Não subestime a sua saúde — a falta de uma simples coenzima no corpo pode custar a vitalidade do paciente."
O manejo do NAD+ representa uma nova fronteira na medicina veterinária integrativa. Compreender que o envelhecimento e a doença crônica são, em essência, crises energéticas celulares permite ao clínico intervir de forma mais assertiva. Cardoso (2025) conclui que "a falta de uma simples coenzima pode trazer vários problemas de saúde e consequências metabólicas" [1].
A transição para a alimentação natural, rica em precursores e livre de antinutrientes processados, é o primeiro e mais importante passo para restaurar a bioenergia. A associação de uma dieta bem formulada com estilo de vida adequado e, quando necessário, o suporte estratégico com precursores, oferece uma abordagem completa para atacar a disfunção mitocondrial na sua raiz.
9 REFERÊNCIAS
1. CARDOSO, D. NAD+: a coenzima que corrige (quase) tudo. [S.l.]: Mundo Hormonal, 2025. 1 vídeo (aprox. 20 min). Disponível em: https://youtu.be/t1NlydtSodc. Acesso em: 16 maio 2026.
2. HARDEN, A.; YOUNG, W. J. The alcoholic ferment of yeast-juice. Proceedings of the Royal Society of London. Series B, v. 78, n. 526, p. 369-375, 1906.
3. IMLAI, S. The NAD World 2.0: the importance of the inter-tissue communication mediated by NAMPT/NAD+/SIRT1 in mammalian aging and longevity control. npj Systems Biology and Applications, v. 2, p. 16018, 2016.
4. YOSHINO, J.; BAUR, J. A.; IMLAI, S. NAD+ intermediates: the biology and therapeutic potential of NMN and NR. Cell Metabolism, v. 27, n. 3, p. 513-528, 2018.
5. CANTO, C.; AUWERX, J. NAD+ as a signaling molecule modulating metabolism. Current Opinion in Cell Biology, v. 24, n. 2, p. 258-265, 2012.
6. VERDIN, E. NAD+ in aging, metabolism, and neurodegeneration. Science, v. 350, n. 6265, p. 1208-1213, 2015.
7. COVARRUBIAS, A. J. et al. NAD+ metabolism and its roles in cellular processes during ageing. Nature Reviews Molecular Cell Biology, v. 22, n. 2, p. 119-141, 2021.
8. KATAYOSHI, C. et al. Nicotinamide mononucleotide (NMN) supplementation improves muscle function in aged mice. Nature Communications, v. 7, p. 13164, 2016.
9. MITSCH, A. et al. NAD+ metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus. Cell Metabolism, v. 22, n. 1, p. 31-53, 2015.
10. NIKIFOROV, A. et al. Pathways and subcellular compartmentation of NAD biosynthesis in human cells. Journal of Biological Chemistry, v. 289, n. 34, p. 23713-23725, 2014.
PETCLUBE INSTITUTE OF INTEGRATIVE VETERINARY MEDICINE
NAD+ AS THE CENTRAL AXIS OF CELLULAR BIOENERGETICS: NATURAL NUTRITIONAL SOURCES, ENERGY DISPOSITION, AND INTEGRATIVE STRATEGIES IN NATURAL FEEDING FOR DOGS AND CATS
"NAD+ is not just a coenzyme — it is the bridge between the food the animal consumes and the energy its cells utilize."
16 de maio de 2026
ABSTRACT
"Without sufficient NAD+, the cell repairs worse, regulates worse, protects worse, and produces energy poorly."
Nicotinamide Adenine Dinucleotide (NAD+) is the central coenzyme of cellular bioenergetics, acting as an indispensable mediator in energy metabolism and the primary activator of sirtuins. This article provides a comprehensive review of the role of NAD+ in the energy disposition of dogs and cats, addressing its physiological decline associated with aging—a process exacerbated by chronic inflammation, oxidative damage, and metabolic stress. We analyze the "three great machines" that consume NAD+: sirtuins (the longevity team), PARPs (DNA repair mechanisms), and the CD38 enzyme (the inflammatory "thief" of NAD+). The study highlights the clinical inefficacy of direct oral NAD+ supplementation due to gastric degradation and molecular size, advocating instead for the superiority of Natural Feeding (NF) as a robust source of bioavailable precursors such as niacin, tryptophan, NMN, and NR. Furthermore, we discuss the nutritional defasage of commercial extruded kibble, where high-temperature processing degrades essential B-complex vitamins. Finally, we explore the therapeutic synergy of NAD+ with mitochondrial peptides SS31 and MOTS-c for mitochondrial repair, offering an integrative framework for managing vitality and longevity in small animal practice.
Keywords: NAD+. Mitochondrial dysfunction. Natural feeding. Sirtuins. Integrative veterinary medicine.
1. INTRODUCTION
"NAD+ is the central molecule of bioenergetics — without it, the mitochondrial plant stops, and the animal pays the price with fatigue, inflammation, and premature aging."
NAD+ (Nicotinamide Adenine Dinucleotide) is an essential coenzyme present in all living cells, serving as a cornerstone for biological existence. As Cardoso (2025) emphasizes, "NAD+ is not a peptide, it is a coenzyme, a central molecule of bioenergetics" [1]. Its primary biological function is to act as a universal electron acceptor in oxidation-reduction (redox) reactions. This role is critical for the catabolism of macronutrients—carbohydrates, lipids, and proteins—enabling their conversion into usable chemical energy in the form of Adenosine Triphosphate (ATP). Without adequate NAD+ concentrations, the biochemical machinery required for cellular survival becomes sluggish, leading to a systemic failure in energy homeostasis.
The historical trajectory of NAD+ research began with its discovery by Harden and Young in 1906 during studies on yeast fermentation [2]. However, its profound significance in animal physiology was later elucidated by Otto Warburg, who identified it as a key component of cellular respiration. In the context of modern bioenergetics, NAD+ functions as a "transmission cable" that connects the metabolic pathways of the cytoplasm and the mitochondrial matrix. By facilitating the transfer of electrons from the Krebs cycle to the oxidative phosphorylation machinery, NAD+ allows for the massive production of ATP required to sustain complex life [3].
In veterinary clinical practice, the "energy disposition" of dogs and cats is a direct reflection of this mitochondrial efficiency. A pet's ability to perform muscle contractions, maintain thermoregulation, execute cognitive tasks, and mount an immune response is entirely dependent on the organism's capacity to extract energy from food and distribute it effectively. All these high-demand functions require ATP, and ATP production is strictly gated by NAD+ availability. As Cardoso (2025) notes, "without sufficient NAD+ in your body, the cell repairs worse, regulates worse, protects worse, and produces energy poorly" [1]. Consequently, NAD+ deficiency manifests clinically as lethargy, exercise intolerance, and a diminished "spark" in geriatric patients.
Natural Feeding (NF) for dogs and cats, characterized by fresh, minimally processed ingredients, offers a significant physiological advantage in maintaining NAD+ levels. Fresh dietary sources such as beef liver, heart, sardines, eggs, and muscle meats are naturally dense in niacin (vitamin B3), tryptophan, riboflavin (B2), and pyridoxine (B6). These are the essential precursors and cofactors required for the three main NAD+ biosynthetic pathways. Unlike synthetic isolates, the complex food matrix found in NF provides these nutrients in highly bioavailable forms, accompanied by synergistic enzymes and minerals that facilitate their conversion into active coenzymes [4].
Conversely, the reliance on commercial extruded kibble presents a significant nutritional challenge. The extrusion process typically involves high temperatures (ranging from 120°C to 180°C) and high pressure, which significantly degrades heat-sensitive B-complex vitamins. Studies indicate that thiamine (B1), riboflavin (B2), and niacin (B3) can suffer losses of up to 60% during processing [5]. Furthermore, the bioavailability of tryptophan—the essential amino acid precursor for the De Novo pathway—is often compromised by the Maillard reaction during kibble production. It is also critical to note that direct oral NAD+ supplementation is largely ineffective; the molecule's size and charge prevent efficient cellular uptake, and it is rapidly degraded by gastric acid [3]. Therefore, providing the correct precursors through a fresh, species-appropriate diet remains the most effective clinical strategy.
The clinical relevance of NAD+ management cannot be overstated. As the veterinary community shifts toward integrative and preventive models, addressing mitochondrial dysfunction becomes paramount. This article aims to review the biochemistry of NAD+, its biosynthesis pathways, and the impact of nutrition on its levels. By understanding the anti-inflammatory and bioenergetic defense mechanisms mediated by this coenzyme, practitioners can better address the root causes of aging and chronic degenerative diseases in small animals, moving beyond mere symptom management toward true metabolic restoration.
2. BIOCHEMISTRY OF NAD+: MECHANISMS AND CELLULAR FUNCTIONS
"NAD+ is not a final product — it is the cable that transmits raw energy into the mitochondria, and without it the cellular plant stalls."
The molecular architecture of NAD+ consists of two nucleotides—nicotinamide and adenine—joined by their phosphate groups. Its primary metabolic utility stems from its ability to cycle between two states: the oxidized form (NAD+) and the reduced form (NADH). This redox cycling is the heartbeat of cellular metabolism. The fundamental reaction is represented as:
NAD++H++2e−⇌NADH
In the mitochondrial matrix, NAD+ acts as a high-affinity electron scavenger during the Krebs cycle (Citric Acid Cycle). As acetyl-CoA is oxidized, NAD+ captures the released electrons, transforming into NADH. This NADH then proceeds to the Electron Transport Chain (ETC) located on the inner mitochondrial membrane. Here, NADH delivers its electron payload to Complex I (NADH dehydrogenase), initiating a flow of electrons that powers the pumping of protons across the membrane. This creates the electrochemical gradient (proton motive force) that drives the ATP synthase "turbine" to generate ATP. Cardoso (2025) clarifies this relationship: "NAD+ is the raw form needed in mitochondria to generate ATP, while NADH delivers energy in a more constant form. For NADH to work, you can't take it alone — you need both together" [1].
Beyond its role in ATP production, NAD+ serves a second, equally vital function as a signaling molecule and substrate for specialized enzymes. The most prominent of these are the sirtuins, a family of NAD+-dependent deacetylases. Cardoso (2025) describes sirtuins as "the longevity team — masters of cellular maintenance and adaptation" [1]. When NAD+ levels are high, sirtuins are activated, triggering a cascade of protective mechanisms. SIRT1, located primarily in the nucleus, coordinates DNA repair and activates PGC-1α, which is the master regulator of mitochondrial biogenesis—the creation of new, healthy mitochondria. SIRT3, the primary mitochondrial sirtuin, works to reduce electron leakage and neutralize reactive oxygen species (ROS), while SIRT6 is essential for telomere maintenance and genomic stability.
However, the availability of NAD+ for these protective sirtuins is constantly threatened by other metabolic "consumers." Cardoso (2025) identifies the "three great NAD+-consuming machines" that dictate the coenzyme's fate [1]. The first are the PARPs (Poly ADP-ribose polymerases). These enzymes function as "DNA mechanics." When they detect genomic damage caused by oxidative stress, toxins, or radiation, they consume massive quantities of NAD+ to facilitate repair. In a state of chronic damage—often seen in sedentary, obese, or poorly nourished pets—PARP activity can become so high that it exhausts the cellular NAD+ pool, leaving nothing for the sirtuins to promote longevity.
The third and perhaps most insidious consumer is CD38, often referred to as the "NAD+ thief." CD38 is an ectoenzyme that degrades NAD+ into nicotinamide and cADPR. Crucially, CD38 expression increases dramatically in response to chronic inflammatory processes. As an animal ages or suffers from metabolic syndrome, systemic levels of pro-inflammatory cytokines such as IL-6 and TNF-α rise, which in turn upregulates CD38. This creates a "sink" for NAD+, where the coenzyme is destroyed faster than it can be synthesized or recycled. This inflammatory depletion of NAD+ is a primary driver of the metabolic decline seen in geriatric dogs and cats [1].
3. NAD+ DECLINE WITH AGING AND CLINICAL IMPLICATIONS
"The NAD+ decline is physiological — aging is, to a large extent, losing the capacity to produce energy."
The aging process in companion animals is inextricably linked to a progressive reduction in systemic NAD+ concentrations. Cardoso (2025) states unequivocally: "the decline is physiological — with aging it is normal for NAD+ levels to fall. The older you get, the less NAD+ you have. And it worsens with inflammation, damage, and metabolic stress" [1]. This decline is not merely a marker of age but a functional cause of the physiological deterioration we observe in senior pets. When NAD+ levels drop below a critical threshold, the cell's ability to maintain homeostasis is compromised, leading to a state of "bioenergetic bankruptcy."
This decline is driven by three primary mechanisms. First, the efficiency of the NAMPT-mediated salvage pathway—the body's primary method for recycling NAD+—diminishes with age. Second, the accumulation of DNA damage over a lifetime leads to chronic overactivation of PARPs, which drain the NAD+ pool. Third, the "inflammaging" phenomenon (chronic, low-grade systemic inflammation) causes a steady increase in CD38 activity, which actively destroys available NAD+ [1, 6]. Together, these factors create a metabolic pincer movement that starves the mitochondria of their essential coenzyme.
The clinical consequences of this decline are most evident in high-energy tissues. In skeletal muscle, low NAD+ contributes to sarcopenia and muscle wasting, as the sirtuin-mediated pathways for muscle protein synthesis and mitochondrial quality control fail. In the liver, NAD+ depletion is a precursor to insulin resistance and hepatic steatosis, as the organ loses its ability to efficiently oxidize fatty acids. In the heart, reduced NAD+ levels are associated with hypertrophic cardiomyopathy and reduced stroke volume, as the myocardium is unable to meet its immense ATP demands [1, 7].
Furthermore, Cardoso (2025) draws a direct connection between NAD+ depletion and "brain rot" or neurodegeneration [1]. The central nervous system is exceptionally sensitive to energy deficits. Sirtuin activation is vital for protecting neurons against the accumulation of toxic proteins, such as beta-amyloid. When NAD+ is low, the brain's resilience to oxidative stress and neuroinflammation collapses. This is often reflected in the elevation of NFL (Neurofilament Light Chain), a biomarker of axonal damage. Clinically, this manifests as Canine Cognitive Dysfunction (CCD), characterized by disorientation, altered sleep-wake cycles, and loss of house training—a tragic "rotting" of the cognitive faculties that can be mitigated by restoring mitochondrial health [1].
4. NAD+ BIOSYNTHESIS PATHWAYS AND NATURAL SOURCES
"The body produces, recycles, and spends NAD+ — and natural feeding is the most powerful tool to sustain this balance."
To maintain adequate NAD+ levels, the organism relies on three distinct biosynthetic pathways. Understanding these pathways allows the clinician to use Natural Feeding as a targeted therapeutic tool.
4.1 Preiss-Handler Pathway
This pathway converts nicotinic acid (a form of niacin) into NAD+ through the enzyme NAPRT. It is a highly efficient route for increasing systemic NAD+ levels. The richest natural sources for this pathway are found in animal tissues, particularly beef liver, heart, kidney, and oily fish like sardines and salmon. Incorporating these organ meats into a pet's diet provides a direct "jolt" of nicotinic acid that the liver can readily convert into active coenzymes.
4.2 Salvage Pathway (Recycling)
The salvage pathway is the most critical route, responsible for recycling over 85% of the body's daily NAD+ needs. It takes nicotinamide (NAM)—a byproduct of NAD+ consumption by sirtuins and PARPs—and converts it back into NAD+ via the rate-limiting enzyme NAMPT. Cardoso (2025) explains: "NAMPT is the enzyme that helps transform used material back into useful NAD+. But with age, this pathway loses strength and recycling drops" [1]. While this pathway is largely internal, its efficiency is enhanced by lifestyle factors such as physical exercise and the avoidance of excessive caloric intake, both of which are easily managed in a Natural Feeding framework.
4.3 De Novo Pathway (Tryptophan)
The De Novo pathway synthesizes NAD+ from the essential amino acid tryptophan. However, this pathway is notoriously inefficient, with only about 1-2% of dietary tryptophan being converted to NAD+ in most mammals. Furthermore, this conversion is "expensive" in terms of cofactors, requiring vitamin B6 (pyridoxine), vitamin B2 (riboflavin), and Iron as obligatory catalysts. Natural sources rich in both tryptophan and these cofactors include eggs, turkey, chicken, and ruminant meats. A deficiency in any of these cofactors—common in highly processed diets—will stall this pathway, leading to metabolic bottlenecks.
4.4 Direct Precursors
While NAD+ itself is not easily absorbed, its intermediates are. Nicotinamide Riboside (NR) is found in trace amounts in fresh milk. Nicotinamide Mononucleotide (NMN) is found in various fresh vegetables such as avocado, broccoli, cabbage, and cucumber [8]. While the concentrations in these foods are lower than in organ meats, they provide a steady, natural supply of precursors that support the salvage pathway without the risks associated with high-dose synthetic isolates.
5. NATURAL FEEDING AS A NUTRITIONAL STRATEGY FOR NAD+ SUPPORT
"Physique is a consequence of good habits — and the mitochondria respond first to the quality of what the animal ingests."
The superiority of Natural Feeding (NF) in supporting NAD+ levels lies in its preservation of micronutrient integrity and bioavailability. Commercial kibble, by its very nature, is a product of extreme processing. The extrusion process (120-180°C) not only destroys a significant portion of the B-vitamin complex but also creates advanced glycation end-products (AGEs) that trigger the very inflammation that activates the "NAD+ thief," CD38 [5]. By transitioning a pet to NF, we remove these pro-inflammatory triggers while simultaneously providing a dense array of bioavailable precursors.
Organ meats, specifically the liver and heart, should be viewed as "mitochondrial superfoods." The heart is an exceptionally rich source of both niacin and Coenzyme Q10, which works alongside NAD+ in the electron transport chain. The liver provides the full spectrum of B-vitamins (B2, B6, B12) and iron necessary for the De Novo pathway. In NF, these nutrients are delivered within a biological matrix that includes fats and proteins that facilitate their absorption, a synergy that is often lost in the "purified" environment of a synthetic supplement added to kibble [9].
Cardoso (2025) emphasizes that "it's useless to use a peptide or coenzyme if the base isn't correct — diet, training, and sleep need to be aligned. Physique is a consequence of good habits" [1]. In the veterinary context, this means that NAD+ support must be part of a holistic lifestyle change. "Training" for a dog involves consistent physical activity and environmental enrichment that stimulates NAMPT activity. "Sleep" involves providing a stress-free environment where the animal can enter deep restorative phases, allowing sirtuins to perform their maintenance work without the interference of high cortisol levels.
For geriatric patients or those with chronic illness, the strategy should be "supplementation via the food matrix." This involves fortifying a fresh diet with specific high-precursor foods. For example, adding small amounts of sardines (rich in niacin and Omega-3s) or lightly steamed broccoli (source of NMN) can provide a steady influx of NAD+ building blocks. This approach is far more sustainable and physiologically sound than attempting to bypass the digestive system with oral NAD+ capsules, which Cardoso (2025) notes are largely ineffective [1].
Ultimately, the goal of NF is to restore the animal's metabolic flexibility. By providing a diet that is low in inflammatory carbohydrates and high in mitochondrial precursors, we lower the activity of CD38 and PARPs while fueling the sirtuins. This shift allows the animal to move from a state of "survival metabolism" to one of "thriving metabolism," where energy is available not just for basic life support, but for repair, play, and cognitive engagement.
6. FORMS OF ENERGY PRODUCTION AND ANTI-INFLAMMATORY DEFENSE OF NAD+
"Without sufficient NAD+, the mitochondrial plant doesn't stop completely, but it loses efficiency and creates chaos in metabolism."
NAD+ is the ultimate mediator between catabolism (breaking down food) and anabolism (building and repairing the body). When NAD+ levels are insufficient, the Krebs cycle slows down, and the Electron Transport Chain (ETC) fails to receive a steady supply of electrons. This leads to a "stalling" of the mitochondrial plant. Cardoso (2025) explains the fallout: "Low NAD+ worsens fat oxidation, worsens energy production, and increases ROS (reactive oxygen species). It creates chaos in metabolism" [1]. This metabolic chaos is characterized by an increase in oxidative stress, which further damages the mitochondria, creating a self-perpetuating cycle of decay.
The anti-inflammatory defense provided by NAD+ is primarily mediated through sirtuin signaling. SIRT1 is a potent inhibitor of NF-κB, the master switch for the body's inflammatory response. By deacetylating NF-κB, SIRT1 prevents the transcription of pro-inflammatory genes, effectively "cooling down" systemic inflammation. SIRT3 protects the mitochondria by deacetylating and activating antioxidant enzymes like SOD2 (Superoxide Dismutase), which neutralizes the ROS generated during ATP production. SIRT6 further modulates the inflammatory response by suppressing TNF-α expression.
Cardoso (2025) warns that "inflammation accelerates NAD+ depletion" [1]. This creates a dangerous feedback loop: chronic inflammation (from poor diet or obesity) activates CD38, which destroys NAD+. The resulting low NAD+ levels mean sirtuins cannot inhibit NF-κB, leading to even more inflammation. This cycle is the "root cause" of many age-related diseases in pets, from osteoarthritis to chronic kidney disease.
Natural Feeding is the most effective way to break this cycle. By providing natural antioxidants—such as the polyphenols found in berries or the sulforaphane in cruciferous vegetables—we directly reduce systemic inflammation. This lowers the demand on CD38, preserving the NAD+ pool for sirtuin-mediated repair. Thus, NF acts as both a "fuel provider" (precursors) and a "shield" (anti-inflammatories) for the animal's bioenergetic system [10].
7. SYNERGY BETWEEN NAD+ AND MITOCHONDRIAL PEPTIDES IN INTEGRATIVE PRACTICE
"NAD+ fuels the mitochondria — SS31 and MOTS-c structure them. Together, they attack the root cause of aging."
The clinical efficacy of NAD+ is heavily dependent on the structural integrity of the mitochondria. The inner mitochondrial membrane contains a unique phospholipid called cardiolipin, which acts as a scaffold for the ETC complexes. Over time, cardiolipin becomes oxidized and damaged, causing the ETC complexes to drift apart and lose efficiency. Cardoso (2025) suggests a synergistic approach: "NAD+ pairs beautifully with MOTS-c and SS31, especially if you're thinking about mitochondrial repair or optimization" [1].
SS31 (Elamipretide) is a peptide that specifically targets and protects cardiolipin, restoring the structural organization of the ETC. When SS31 "fixes the machinery," the NAD+ provided through diet or precursors can be used much more efficiently to produce ATP. MOTS-c, a mitochondrial-derived peptide, acts as a metabolic regulator that enhances glucose metabolism and increases the expression of NAMPT, thereby boosting the NAD+ salvage pathway. Together, these interventions address both the "fuel" (NAD+) and the "engine" (mitochondrial structure).
Cardoso (2025) also clarifies the distinction between NAD+ and NADH in therapy: "NAD+ is the raw form to give that jolt to mitochondria to refeed them and restart energy production. NADH provides a more consistent, constant energy balance. But it only does this if it has energy. Without NAD+, you don't have enough energy in the mitochondria. That's why some people take NADH thinking they'll improve energy and they improve nothing — you need both together" [1]. In a clinical setting, focusing on NAD+ precursors through Natural Feeding ensures that the "raw form" is always available to maintain this balance.
The timeline for observing significant clinical results in mitochondrial restoration is typically 3 to 6 months. As Cardoso (2025) notes, "it takes months to change habits and see mitochondrial restoration" [1]. This requires patience from both the clinician and the pet owner, emphasizing that true health is a long-term investment in cellular bioenergetics rather than a quick fix.
8. CONCLUSION
"Don't underestimate your health — the lack of a simple coenzyme in the body can cost the patient's vitality."
The management of NAD+ levels represents a paradigm shift in integrative veterinary medicine. As Cardoso (2025) concludes, "the anti-aging process is linked to countless factors. The lack of a simple coenzyme can bring several health problems and metabolic consequences" [1]. By recognizing that aging and chronic disease are essentially energy crises at the cellular level, veterinarians can move toward more sophisticated and effective interventions.
The transition to Natural Feeding is the foundational step in this process. By providing a diet rich in bioavailable precursors and cofactors, we support the body's natural biosynthetic and recycling pathways. This nutritional base, combined with a lifestyle that promotes physical activity and reduces inflammatory stress, creates the ideal environment for NAD+ to perform its dual roles in energy production and cellular defense.
In conclusion, addressing mitochondrial dysfunction through the NAD+ axis offers a powerful tool for restoring vitality in our patients. Whether through the strategic use of organ meats in Natural Feeding or the synergistic application of mitochondrial peptides, the goal remains the same: to ensure that the "mitochondrial plant" of our dogs and cats has the fuel and the structure it needs to thrive. By attacking the root cause of biological failure, we provide our patients not just with more years of life, but with more life in their years.
REFERENCES
1. CARDOSO, D. NAD+: the coenzyme that fixes (almost) everything. [S.l.]: Mundo Hormonal, 2025. 1 video (20 min). Available at: https://youtu.be/t1NlydtSodc. Accessed: May 16, 2026.
2. HARDEN, A.; YOUNG, W. J. The alcoholic ferment of yeast-juice. Proceedings of the Royal Society of London. Series B, v. 78, n. 526, p. 369-375, 1906.
3. IMLAI, S. The NAD World 2.0: the importance of the inter-tissue communication mediated by NAMPT/NAD+/SIRT1 in mammalian aging and longevity control. npj Systems Biology and Applications, v. 2, p. 16018, 2016.
4. YOSHINO, J.; BAUR, J. A.; IMLAI, S. NAD+ intermediates: the biology and therapeutic potential of NMN and NR. Cell Metabolism, v. 27, n. 3, p. 513-528, 2018.
5. CANTO, C.; AUWERX, J. NAD+ as a signaling molecule modulating metabolism. Current Opinion in Cell Biology, v. 24, n. 2, p. 258-265, 2012.
6. VERDIN, E. NAD+ in aging, metabolism, and neurodegeneration. Science, v. 350, n. 6265, p. 1208-1213, 2015.
7. COVARRUBIAS, A. J. et al. NAD+ metabolism and its roles in cellular processes during ageing. Nature Reviews Molecular Cell Biology, v. 22, n. 2, p. 119-141, 2021.
8. KATAYOSHI, C. et al. Nicotinamide mononucleotide (NMN) supplementation improves muscle function in aged mice. Nature Communications, v. 7, p. 13164, 2016.
9. MITSCH, A. et al. NAD+ metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus. Cell Metabolism, v. 22, n. 1, p. 31-53, 2015.
10. NIKIFOROV, A. et al. Pathways and subcellular compartmentation of NAD biosynthesis in human cells. Journal of Biological Chemistry, v. 289, n. 34, p. 23713-23725, 2014.
PETCLUBE 整合兽医学研究所
NAD+ 作为细胞生物能量学的核心轴:天然营养来源、能量分配及犬猫生食喂养中的整合策略
"NAD+ 不仅仅是一种辅酶——它是动物摄入的食物与其细胞利用的能量之间的桥梁。"
2026年5月16日
摘要 (ABSTRACT)
"没有足够的NAD+,细胞修复更差、调节更差、保护更差,能量产生也差。"
烟酰胺腺嘌呤二核苷酸(NAD+)是细胞生物能量学的核心辅酶,在能量代谢和去乙酰化酶(Sirtuins)激活中发挥着不可替代的作用。本文综述了 NAD+ 在犬猫能量分配中的关键地位,探讨了随衰老而出现的生理性下降及其临床影响。重点分析了消耗 NAD+ 的“三大机器”:去乙酰化酶(长寿团队)、PARPs(DNA 修复)和 CD38 酶(炎症引起的降解)。研究表明,直接口服 NAD+ 补充剂因胃肠道降解而效果不佳,而生食喂养(Natural Feeding)作为生物可利用前体(如烟酸、色氨酸、NMN 和 NR)的来源具有显著优势。相比之下,商业挤压粮由于高温加工导致 B 族维生素严重匮乏。最后,本文探讨了 NAD+ 与线粒体肽(SS31 和 MOTS-c)在修复线粒体功能方面的协同作用,为整合兽医学临床实践中促进动物健康和长寿提供了系统性策略。
关键词:NAD+;线粒体功能障碍;生食喂养;去乙酰化酶;整合兽医学
1 引言 (INTRODUCTION)
"NAD+ 是生物能量学的核心分子——没有它,线粒体工厂就会停工,动物就会付出疲劳、炎症和过早衰老的代价。"
NAD+(烟酰胺腺嘌呤二核苷酸)是存在于所有活细胞中的必需辅酶,是生命维持的基石。正如 Cardoso (2025) 所强调的,“NAD+ 不是一种肽,它是一种辅酶,是生物能量学的核心分子” [1]。其主要功能是在氧化还原反应中充当电子受体,使大分子营养物质能够转化为以三磷酸腺苷(ATP)形式存在的可用化学能量。在兽医临床中,NAD+ 的水平直接决定了患宠的生命活力和抗病能力。
从历史角度看,NAD+ 由 Harden 和 Young 于 1906 年发现,随后由 Otto Warburg 阐明其作为细胞呼吸关键成分的作用 [2]。它在生化上充当“传输电缆”,将克雷布斯循环(三羧酸循环)与氧化磷酸化连接起来 [3]。这种连接确保了电子流能够高效地通过线粒体呼吸链,从而产生维持生命所需的动力。对于犬猫等高代谢率的肉食动物而言,这一过程的完整性至关重要。
宠物进行肌肉收缩、维持体温、执行认知任务以及产生免疫反应的能力完全取决于 ATP 的产生,而 ATP 的产生受到 NAD+ 可用性的严格限制。Cardoso (2025) 指出,“没有足够的 NAD+,细胞修复更差、调节更差、保护更差,能量产生也差” [1]。临床上,NAD+ 水平低下的动物常表现为嗜睡、运动耐力下降和康复缓慢,这些症状往往被错误地归因于“正常”衰老。
基于新鲜食材(如牛肝、心脏、沙丁鱼、鸡蛋)的生食喂养在维持 NAD+ 水平方面具有显著优势。这些食材富含烟酸(维生素 B3)、色氨酸、核黄素(B2)和吡哆醇(B6) [4]。生食喂养的复杂食物基质有利于这些前体物质的协同吸收,尊重肉食动物的消化生理,并优化内源性生物合成。这种营养策略不仅提供了原料,还提供了维持代谢平衡所需的辅助因子。
相比之下,商业挤压粮面临严重的热加工挑战。在 120-180°C 的挤压过程中,30-60% 的 B 族维生素会发生降解,并降低色氨酸的生物利用度。此外,研究证实直接口服 NAD+ 会因胃酸和消化酶的作用而失效 [3, 5]。因此,对于追求卓越性能和长寿的患宠,转向生食喂养或精准的营养干预已成为临床必然。本文旨在综述 NAD+ 的生化机制、生物合成途径及其在整合兽医学中的应用前景。
2 NAD+ 的生物化学:机制与细胞功能
"NAD+ 不是最终产物——它是将原始能量传输到线粒体的电缆,没有它,细胞工厂就会停工。"
NAD+ 的分子结构由两个通过磷酸基团连接的核苷酸组成。其代谢动力学基于氧化态(NAD+)和还原态(NADH)之间的交替。在克雷布斯循环中,NAD+ 捕获来自碳水化合物和脂肪氧化的电子。Cardoso (2025) 解释说,“NAD+ 是线粒体产生 ATP 所需的原始形式,而 NADH 则以更恒定的形式传递能量。你需要两者共同作用” [1]。这种平衡对于线粒体呼吸链维持产生 ATP 所需的电化学梯度至关重要。
除了能量产生,NAD+ 还具有关键的信号传导功能,即激活去乙酰化酶(Sirtuins)。Cardoso (2025) 将这些蛋白质描述为“长寿团队——细胞维护和适应的大师” [1]。在兽医实践中,三种异构体尤为重要:SIRT1 作用于细胞核,促进 DNA 修复并激活 PGC-1α(线粒体生物发生的调节器);SIRT3 位于线粒体基质,负责减少氧化应激;SIRT6 则对端粒保护和基因组稳定性至关重要。
然而,NAD+ 的可用性受到细胞内“三大机器”竞争的影响:去乙酰化酶、PARPs 和 CD38 [1]。PARPs(聚 ADP-核糖聚合酶)被形象地称为“DNA 修理机”,当它们检测到基因组损伤时,会消耗大量 NAD+ 进行修复。如果损伤持续存在,这种修复过程可能会耗尽细胞的代谢系统。CD38 酶则被称为“NAD+ 小偷”,它随着炎症水平的升高而增加,直接降解 NAD+,导致能量匮乏。
3 NAD+ 随衰老下降及临床意义
"NAD+ 的下降是生理性的——衰老在很大程度上就是失去产生能量的能力。"
随着犬猫步入老年,其体内的 NAD+ 水平呈现出明显的下降趋势。Cardoso (2025) 强调,“这种下降是生理性的——随着衰老,NAD+ 水平下降是正常的。但炎症、损伤和代谢压力会加剧这一过程” [1]。这种下降并非单一因素引起,而是由合成减少和消耗增加共同导致的。具体而言,负责回收 NAD+ 的 NAMPT 途径随着年龄增长而失去强度,而 CD38 的表达却因慢性炎症而显著上调 [6]。
NAD+ 匮乏对高能耗组织的影响最为严重。在肌肉组织中,这表现为肌肉减少症(Sarcopenia)和收缩力下降;在肝脏中,则导致脂肪氧化受损和胰岛素抵抗。对于老年犬猫,最令人担忧的是神经系统的影响。Cardoso 将低 NAD+ 状态与“大脑腐蚀”(Brain Rot)和神经退行性变联系起来,指出这与 β-淀粉样蛋白的积累和神经丝轻链(NFL)这一神经损伤标志物的升高密切相关 [1]。这种生化环境为认知功能障碍综合征(CDS)埋下了伏笔。
4 NAD+ 生物合成途径与天然来源
"身体产生、回收和消耗 NAD+——而生食喂养是维持这种平衡最强大的工具。"
4.1 Preiss-Handler 途径
该途径利用烟酸(维生素 B3)合成 NAD+。在生食喂养中,牛肝、心脏、肾脏以及沙丁鱼和三文鱼是烟酸的极佳来源。这些食材不仅提供前体,还含有促进转化的酶辅助因子。
4.2 回收途径 (Salvage Pathway)
这是细胞维持 NAD+ 水平的主要方式,占每日产量的 85% 以上。Cardoso (2025) 解释道,“NAMPT 酶帮助将使用过的材料重新转化为有用的 NAD+。随着年龄增长,这一途径会失去力量” [1]。通过摄入富含核苷酸的组织(如胸腺和脾脏),可以为该途径提供底物支持。
4.3 从头合成途径 (De Novo Pathway)
该途径将色氨酸转化为 NAD+,但效率极低(仅约 1-2%)。它需要维生素 B6、B2 和铁作为辅助因子。鸡蛋、火鸡肉和瘦肉是色氨酸的主要来源,但必须确保饮食中微量元素的平衡以维持此途径的运转。
4.4 直接前体 (Direct Precursors)
烟酰胺核苷(NR)和烟酰胺单核苷酸(NMN)是 NAD+ 的直接前体。虽然在自然界中含量较少,但仍存在于牛奶(NR)、鳄梨、西兰花、卷心菜和黄瓜(NMN)中 [8]。在生食配餐中加入少量这些植物性食材,可以作为前体的补充来源。
5 生食喂养作为支持 NAD+ 的营养策略
"体型是好习惯的结果——而线粒体首先对动物摄入的食物质量做出反应。"
生食喂养(Natural Feeding)在支持 NAD+ 代谢方面的优越性源于其微量营养素的完整性。Cardoso (2025) 提醒兽医和宠主,“如果基础不正确,使用肽或辅酶是徒劳的——饮食、训练和睡眠需要保持一致。体型是好习惯的结果” [1]。对于犬猫而言,这意味着必须摆脱高度加工食品的束缚。器官肉被视为“线粒体超级食物”,因为它们浓缩了合成 NAD+ 所需的所有 B 族维生素和辅酶 Q10。
对于老年病例,通过“食物基质补充”是黄金策略。相比于分离的合成补充剂,食物中的营养素以生物可利用的形式存在,并伴有天然抗氧化剂,能有效抑制 CD38 的过度激活。临床观察表明,转为生食喂养的动物在 3 到 6 个月内表现出明显的活力提升和皮毛质量改善,这正是线粒体功能恢复的外部表现。
6 NAD+ 的能量产生形式与抗炎防御
"没有足够的 NAD+,线粒体工厂不会完全停止,但会失去效率并在代谢中造成混乱。"
NAD+ 的缺乏会引发代谢混乱。Cardoso (2025) 指出,“低 NAD+ 会恶化脂肪氧化,恶化能量产生,并增加活性氧(ROS)。它在代谢中制造混乱” [1]。这种混乱不仅表现为能量不足,还表现为促炎环境的形成。当 NAD+ 水平下降时,Sirtuins 无法抑制 NF-κB 等促炎转录因子,导致 IL-6、TNF-α 和 C 反应蛋白(CRP)等炎症标志物升高。
这种炎症状态会进一步激活 CD38,形成一个恶性循环:炎症 → CD38 增加 → NAD+ 耗竭 → 炎症加剧。生食喂养通过提供天然抗炎多酚和充足的 NAD+ 前体,能够有效打破这一循环 [10]。维持高水平的 NAD+ 相当于为细胞安装了一个强大的抗炎屏障,保护线粒体免受氧化损伤。
7 NAD+ 与线粒体肽在整合实践中的协同作用
"NAD+ 为线粒体提供燃料——SS31 和 MOTS-c 构建它们。它们共同攻击衰老的根本原因。"
在高级整合医疗中,NAD+ 常与线粒体肽协同使用。Cardoso (2025) 认为,“NAD+ 与 MOTS-c 和 SS31 完美搭配,用于线粒体修复” [1]。SS31 能够保护线粒体内膜的心磷脂,防止电子泄漏;而 MOTS-c 则调节葡萄糖代谢并刺激 NAMPT 途径。这种组合不仅提供了“燃料”(NAD+),还修复了“发动机”的结构(SS31)。
关于 NAD+ 与 NADH 的区别,Cardoso 提供了清晰的临床指导:“NAD+ 是原始形式,给线粒体带来冲击。NADH 提供恒定能量,但只有在有能量的情况下才起作用——你需要两者结合” [1]。在治疗方案中,通常建议持续使用 3 到 6 个月,并配合对齐的饮食和适当的体力活动,以实现深层的代谢重塑。
8 结论 (CONCLUSION)
"不要低估你的健康——体内缺少一种简单的辅酶就可能危及患者的活力。"
NAD+ 的管理代表了整合兽医学的新前沿。它不仅是一个营养问题,更是生物能量学的核心干预点。Cardoso (2025) 总结道,“抗衰老过程与无数因素有关。缺少一种简单的辅酶会带来多种健康问题和代谢后果” [1]。通过生食喂养提供高质量的前体,结合对线粒体健康的精准支持,兽医师可以从根本上改善犬猫的生活质量。
未来的临床实践应更加关注线粒体健康标志物,并将 NAD+ 轴的优化作为老年病学和慢性病管理的基础步骤。从加工粮向生食喂养的转变,不仅是饮食的改变,更是为细胞重新注入生命能量的基石。
参考文献 (REFERENCES)
1. CARDOSO, D. **NAD+: the coenzyme that fixes (almost) everything**. [S.l.]: Mundo Hormonal, 2025. Available at: https://youtu.be/t1NlydtSodc. Accessed: May 16, 2026.
2. HARDEN, A.; YOUNG, W. J. The alcoholic ferment of yeast-juice. **Proceedings of the Royal Society of London. Series B**, v. 78, n. 526, p. 369-375, 1906.
3. IMLAI, S. The NAD World 2.0. **npj Systems Biology and Applications**, v. 2, p. 16018, 2016.
4. YOSHINO, J.; BAUR, J. A.; IMLAI, S. NAD+ intermediates. **Cell Metabolism**, v. 27, n. 3, p. 513-528, 2018.
5. CANTO, C.; AUWERX, J. NAD+ as a signaling molecule. **Current Opinion in Cell Biology**, v. 24, n. 2, p. 258-265, 2012.
6. VERDIN, E. NAD+ in aging, metabolism, and neurodegeneration. **Science**, v. 350, n. 6265, p. 1208-1213, 2015.
7. COVARRUBIAS, A. J. et al. NAD+ metabolism in ageing. **Nature Reviews Molecular Cell Biology**, v. 22, n. 2, p. 119-141, 2021.
8. KATAYOSHI, C. et al. NMN supplementation improves muscle function in aged mice. **Nature Communications**, v. 7, p. 13164, 2016.
9. MITSCH, A. et al. NAD+ metabolism and energy homeostasis. **Cell Metabolism**, v. 22, n. 1, p. 31-53, 2015.
10. NIKIFOROV, A. et al. Pathways of NAD biosynthesis in human cells. **Journal of Biological Chemistry**, v. 289, n. 34, p. 23713-23725, 2014.
*本文由 Petclube 整合兽医学研究所于 2026年5月16日 编写。所含信息旨在为兽医专业人员提供学术参考。*
מכון פטקלוב לרפואה וטרינרית אינטגרטיבית
NAD+ כציר המרכזי של הביו-אנרגטיקה התאית: מקורות תזונתיים טבעיים, חלוקת אנרגיה ואסטרטגיות אינטגרטיביות בתזונה טבעית לכלבים וחתולים
"NAD+ הוא לא רק קו-אנזים — הוא הגשר בין המזון שהחיה צורכת לבין האנרגיה שהתאים שלה מנצלים."
16 במאי 2026
מחברים: ד"ר קלאודיו אמיקטי ג'וניור – וטרינר אינטגרטיבי (CRMV-SP 75.404 VT, MAPA 00129461/2025), מומחה בתזונה קלינית, רפואה קנבינואידית ותזונה טבעית. ד"ר גבריאל אמיקטי – וטרינר (CRMV-SP 45.592 VT), מומחה באורתופדיה וניתוחי חיות קטנות.
מוסד: מכון פטקלוב לרפואה וטרינרית אינטגרטיבית, סאו פאולו, ברזיל.
תקציר (ABSTRACT)
"ללא מספיק NAD+, התא מתקן גרוע יותר, מווסת גרוע יותר, מגן גרוע יותר ומייצר אנרגיה בצורה לקויה."
ניקוטינאמיד אדנין דינוקלאוטיד (NAD+) מהווה קו-אנזים מרכזי בביו-אנרגטיקה התאית, ומשמש כמתווך חיוני במטבוליזם האנרגטי ובהפעלת סירטואינים. מאמר זה סוקר את תפקידו של NAD+ בחלוקת האנרגיה של כלבים וחתולים, תוך התייחסות לירידה הפיזיולוגית שלו עם הגיל והשפעת הלחץ המטבולי. נדונות "שלוש המכונות הגדולות" הצורכות NAD+: סירטואינים (אריכות ימים), PARPs (תיקון DNA) והאנזים CD38 (פירוק דלקתי). הניתוח מדגיש את חוסר היעילות של תוספי תזונה דרך הפה ואת העליונות של תזונה טבעית (AN) כמקור למבשרי NAD+ בעלי זמינות ביולוגית גבוהה, כגון חומצה ניקוטינית, טריפטופן ונוקלאוטידים. לעומת זאת, מזון מסחרי מעובד (Kibble) מציג חסר תזונתי עקב הרס תרמי של ויטמינים מקבוצת B. לבסוף, נבחנת הסינרגיה של NAD+ עם פפטידים מיטוכונדריאליים (SS31 ו-MOTS-c) בתיקון התפקוד המיטוכונדריאלי, המציעה גישה אינטגרטיבית לקידום בריאות ואריכות ימים ברפואת חיות קטנות.
מילות מפתח: NAD+; תפקוד מיטוכונדריאלי; תזונה טבעית; סירטואינים; רפואה וטרינרית אינטגרטיבית
1 מבוא (INTRODUCTION)
"NAD+ הוא המולקולה המרכזית של הביו-אנרגטיקה — בלעדיה, מפעל המיטוכונדריה נעצר, והחיה משלמת את המחיר בעייפות, דלקת והזדקנות מואצת."
NAD+ (ניקוטינאמיד אדנין דינוקלאוטיד) הוא קו-אנזים חיוני הקיים בכל התאים החיים וממלא תפקיד שאין לו תחליף בהישרדות התאית. כפי שמדגיש קרדוסו (2025), "NAD+ אינו פפטיד, הוא קו-אנזים, מולקולה מרכזית של ביו-אנרגטיקה" [1]. תפקידו העיקרי הוא לשמש כקולט אלקטרונים בתגובות חמצון-חיזור, מה שמאפשר המרה של אבות מזון לאנרגיה כימית זמינה בצורת ATP.
הקו-אנזים התגלה לראשונה על ידי הארדן ויאנג בשנת 1906, ומאוחר יותר הובהר תפקידו על ידי אוטו ורבורג כמרכיב מפתח בנשימה התאית [2]. הוא מתפקד כ"כבל העברה" המחבר את מחזור קרבס לזרחון חמצוני בתוך המיטוכונדריה [3]. ללא זמינות נאותה של מולקולה זו, זרם האלקטרונים נפסק, מה שמוביל לקריסה אנרגטית תאית.
יכולתו של בעל חיים לבצע התכווצויות שרירים, לשמור על ויסות חום, לבצע משימות קוגניטיביות ולהפעיל תגובה חיסונית תלויה לחלוטין בייצור ATP, המוגבל על ידי זמינות ה-NAD+. כפי שציין קרדוסו (2025), "ללא מספיק NAD+, התא מתקן גרוע יותר, מווסת גרוע יותר, מגן גרוע יותר ומייצר אנרגיה בצורה לקויה" [1]. חוסר זה מתבטא קלינית בעייפות כרונית, עמידות לאינסולין והתדרדרות קוגניטיבית.
תזונה טבעית (Natural Feeding) המבוססת על רכיבים טריים כגון כבד בקר, לב, סרדינים וביצים מציעה יתרונות משמעותיים בשמירה על רמות NAD+ דרך אספקת חומצה ניקוטינית, טריפטופן, ריבופלאבין ופירידוקסין [4]. המטריצה התזונתית המורכבת של מזון טרי מאפשרת ספיגה סינרגטית של מבשרים אלו, תוך כיבוד הפיזיולוגיה של טורפים.
לעומת זאת, מזון מסחרי יבש (Kibble) עובר תהליכי אקסטרוזיה בטמפרטורות של 120-180 מעלות צלזיוס, מה שגורם לפירוק של 30-60% מוויטמיני B ומפחית את הזמינות הביולוגית של חומצות אמינו חיוניות. יתרה מכך, מתן NAD+ ישיר דרך הפה אינו יעיל עקב פירוק במערכת העיכול, מה שהופך את האסטרטגיה התזונתית והזרקתית לחיונית [3, 5].
מאמר זה סוקר את הביוכימיה של NAD+, מסלולי הביוסינתזה שלו, מקורות טבעיים ומנגנוני הגנה אנטי-דלקתיים. המטרה היא לספק לווטרינר האינטגרטיבי כלים מדעיים לניהול מטבולי מתקדם של מטופלים כלבים וחתולים.
2 הביוכימיה של NAD+: מנגנונים ותפקודים תאיים
"NAD+ אינו תוצר סופי — הוא הכבל שמעביר אנרגיה גולמית לתוך המיטוכונדריה, ובלעדיו מפעל התא נעצר."
המבנה המולקולרי של NAD+ מורכב משני נוקלאוטידים המחוברים באמצעות קבוצות פוספט. הדינמיקה המטבולית שלו מבוססת על המעבר בין הצורה המחומצנת (NAD+) לצורה המחוזרת (NADH). קרדוסו (2025) מסביר כי "NAD+ הוא הצורה הגולמית הדרושה במיטוכונדריה לייצור ATP, בעוד NADH מספק אנרגיה בצורה קבועה יותר. יש צורך בשניהם יחד" [1].
בתוך המיטוכונדריה, NAD+ פועל כקופקטור חיוני לאנזימים דהידרוגנאזות במחזור קרבס. הוא אוסף אלקטרונים ופרוטונים והופך ל-NADH, אשר לאחר מכן מעביר את המטען לשרשרת העברת האלקטרונים. תהליך זה יוצר את המפל האלקטרוכימי המניע את ה-ATP סינתאז. ללא יחס תקין בין NAD+ ל-NADH, המטבוליזם התאי עובר למסלולים אנאירוביים פחות יעילים, מה שמוביל להצטברות לקטט ועקה חמצונית.
מעבר לתפקידו בייצור אנרגיה, NAD+ משמש כמולקולת איתות קריטית להפעלת סירטואינים. קרדוסו מתאר אותם כ"צוות אריכות הימים — מאסטרים של תחזוקה והסתגלות תאית" [1]. אלו הם אנזימים תלויי NAD+ המווסתים את בריאות התא:
- SIRT1: פועל בגרעין התא, מקדם תיקון DNA ומפעיל את PGC-1α, המווסת המרכזי של יצירת מיטוכונדריות חדשות.
- SIRT3: ממוקם במיטוכונדריה, אחראי על הפחתת רדיקלים חופשיים (ROS) ואופטימיזציה של חמצון חומצות שומן.
- SIRT6: חיוני להגנה על הטלומרים ויציבות הגנום.
קרדוסו (2025) מגדיר את "שלוש המכונות הגדולות" הצורכות NAD+: סירטואינים, PARPs ו-CD38 [1]. ה-PARPs (פולי ADP-ריבוז פולימראזות) מתפקדים כ"מכונאי ה-DNA". כאשר יש נזק גנטי רב עקב קרינה או רעלים, ה-PARPs נכנסים לפעולה אינטנסיבית ועלולים "לרוקן" את מאגרי ה-NAD+ של התא, מה שמוביל למוות תאי (אפופטוזיס) עקב מחסור באנרגיה.
האנזים השלישי, CD38, נחשב ל"גנב ה-NAD+". הוא נמצא על פני השטח של תאי מערכת החיסון ופעילותו עולה משמעותית במצבי דלקת כרונית. ככל שבעל החיים מזדקן וצובר דלקת מערכתית, ה-CD38 מפרק יותר NAD+, מה שמותיר פחות משאבים לייצור אנרגיה ותיקון תאי [1, 6].
3 ירידת NAD+ עם ההזדקנות והשלכות קליניות
"ירידת NAD+ היא פיזיולוגית — ההזדקנות היא, במידה רבה, אובדן היכולת לייצר אנרגיה."
הירידה ברמות ה-NAD+ עם הגיל היא תופעה מתועדת היטב ביונקים. קרדוסו (2025) מציין כי "הירידה היא פיזיולוגית — עם ההזדקנות זה נורמלי שרמות ה-NAD+ ירדו. זה מחמיר עם דלקת, נזק ולחץ מטבולי" [1]. בקרב כלבים וחתולים, ירידה זו מתחילה כבר בגיל העמידה ותורמת ישירות להופעת מחלות ניווניות.
קיימים שלושה מנגנונים עיקריים לירידה זו: (1) ירידה ביעילות מסלול המיחזור (NAMPT), (2) צריכה מוגברת על ידי PARPs עקב נזק מצטבר ל-DNA, ו-(3) עלייה בפירוק על ידי CD38 עקב דלקת כרונית בדרגה נמוכה (Inflammaging) [1, 7].
הרקמות המושפעות ביותר הן אלו בעלות דרישה אנרגטית גבוהה:
- שרירים: מחסור ב-NAD+ מוביל לסרקופניה (אובדן מסת שריר) וחולשה, הנפוצה מאוד בכלבים קשישים.
- מערכת העצבים: המוח צורך כ-20% מהאנרגיה של הגוף. ירידה ב-NAD+ קשורה ל"ריקבון מוחי" (Brain rot) ותפקוד קוגניטיבי לקוי. קרדוסו מקשר רמות נמוכות של NAD+ להצטברות בטא-עמילואיד ולעלייה במדד NFL (Neurofilament Light chain), המעיד על נזק עצבי [1].
- כבד ומטבוליזם: ירידה ב-NAD+ פוגעת בחמצון שומנים ומובילה לעמידות לאינסולין והשמנת יתר.
קלינית, הווטרינר יבחין בבעל חיים ש"איבד את החיוניות שלו". הכלב כבר לא רוצה לשחק, החתול מפסיק לקפוץ למקומות גבוהים, וישנה ירידה כללית ביכולת ההתאוששות ממחלות. מצב זה אינו רק "זקנה", אלא כשל ביו-אנרגטי שניתן לטיפול באמצעות אסטרטגיות תזונתיות ואינטגרטיביות.
4 מסלולי ביוסינתזה של NAD+ ומקורות טבעיים
"הגוף מייצר, ממחזר וצורך NAD+ — ותזונה טבעית היא הכלי החזק ביותר לקיים איזון זה."
הגוף הווטרינרי משתמש בארבעה מסלולים עיקריים לשמירה על רמות NAD+:
4.1 מסלול פרייס-הנדלר (Preiss-Handler)
מסלול זה משתמש בחומצה ניקוטינית (ויטמין B3) כדי לייצר NAD+. המקורות הטבעיים הטובים ביותר הם איברים פנימיים: כבד בקר, לב וכליות. דגים כמו סרדינים וסלמון הם גם מקורות מצוינים. תזונה טבעית עשירה באיברים אלו מבטיחה אספקה קבועה של חומצה ניקוטינית [4].
4.2 מסלול המיחזור (Salvage Pathway)
זהו המסלול החשוב ביותר, הממחזר מעל 85% מה-NAD+ היומי. האנזים NAMPT הופך ניקוטינאמיד (NAM) בחזרה ל-NMN ולאחר מכן ל-NAD+. קרדוסו (2025) מדגיש כי "NAMPT עוזר להפוך חומר משומש בחזרה ל-NAD+ שימושי. עם הגיל, המסלול הזה מאבד מכוחו" [1]. פעילות גופנית ותזונה נכונה יכולים להמריץ מסלול זה.
4.3 מסלול דה נובו (De Novo Pathway)
ייצור NAD+ מחומצת האמינו טריפטופן. מסלול זה אינו יעיל במיוחד (יחס של 60:1) ודורש נוכחות של ויטמין B6, B2 וברזל כקופקטורים. מקורות לטריפטופן כוללים ביצים, הודו ובשר בקר. מחסור בוויטמיני B בתזונה (הנפוץ במזון יבש מעובד) פוגע קשות במסלול זה.
4.4 תוצרי ביניים ישירים (Direct Precursors)
מבשרים כמו NR (ניקוטינאמיד ריבוזיד) ו-NMN (ניקוטינאמיד מונו-נוקלאוטיד) נמצאים בכמויות קטנות במזונות כמו חלב (NR), אבוקדו, ברוקולי ומלפפונים (NMN) [8]. למרות שהכמויות קטנות, צריכה קבועה דרך תזונה טבעית תורמת למאגר התאי.
5 תזונה טבעית כאסטרטגיה תזונתית לתמיכה ב-NAD+
"מבנה גוף הוא תוצאה של הרגלים טובים — והמיטוכונדריה מגיבה תחילה לאיכות המזון שהחיה צורכת."
המעבר למזון טרי (AN) הוא הצעד הראשון והחשוב ביותר בשיקום הביו-אנרגטיקה של המטופל. מזון מסחרי יבש עובר תהליך של אקסטרוזיה שפוגע בשלמות המיקרו-נוטריאנטים. לעומת זאת, תזונה טבעית מאוזנת מספקת את כל הקופקטורים הדרושים (מגנזיום, ויטמיני B, אבץ) לסנתזה של NAD+.
קרדוסו (2025) טוען כי "זה חסר תועלת להשתמש בפפטיד או בקו-אנזים אם הבסיס אינו נכון — תזונה, אימון ושינה צריכים להיות מתואמים. מבנה גוף הוא תוצאה של הרגלים טובים" [1]. ברפואה וטרינרית, "אימון" מתרגם לגירוי פיזי ומנטלי מותאם לגיל, ו"שינה" לסביבה נטולת מתח.
שימוש באיברים פנימיים (Offal) בתזונה טבעית הוא קריטי. הכבד אינו רק מסנן רעלים, אלא מחסן של ויטמינים מקבוצת B ומינרלים. הלב עשיר ב-CoQ10 וטאורין, הפועלים בסינרגיה עם NAD+ בשרשרת העברת האלקטרונים. עבור מטופלים גריאטריים, "תיסוף דרך מטריצת המזון" היא האסטרטגיה המנצחת, שכן היא מונעת את הפירוק המהיר שקורה בתוספים סינתטיים מבודדים.
6 צורות ייצור אנרגיה והגנה אנטי-דלקתית של NAD+
"ללא מספיק NAD+, מפעל המיטוכונדריה לא נעצר לגמרי, אבל מאבד מיעילות ויוצר כאוס במטבוליזם."
כאשר רמות ה-NAD+ נמוכות, המיטוכונדריה מייצרת יותר רדיקלים חופשיים (ROS) ופחות ATP. קרדוסו (2025) מסביר: "NAD+ נמוך מחמיר את חמצון השומן, מחמיר את ייצור האנרגיה ומגביר את ה-ROS. זה יוצר כאוס במטבוליזם" [1]. חוסר יעילות זה מוביל לדלקת תאית.
NAD+ פועל כנוגד דלקת עוצמתי דרך מספר מסלולים:
- עיכוב NF-κB: SIRT1, המופעל על ידי NAD+, מעכב את גורם השעתוק NF-κB, שהוא המנוע העיקרי של דלקת בגוף.
- דיכוי ציטוקינים: SIRT6 מדכא את הייצור של TNF-α וציטוקינים דלקתיים אחרים.
- ניקוי רעלים חמצוני: SIRT3 מפעיל אנזימים נוגדי חמצון בתוך המיטוכונדריה.
נוצר מעגל קסמים שלילי: דלקת מעלה את רמת ה-CD38, שמוריד את ה-NAD+, מה שמוביל ליותר דלקת. תזונה טבעית עשירה בנוגדי חמצון ופיטו-נוטריאנטים עוזרת לשבור מעגל זה על ידי הפחתת העומס הדלקתי הראשוני [10].
7 סינרגיה בין NAD+ לפפטידים מיטוכונדריאליים בפרקטיקה אינטגרטיבית
"NAD+ מתדלק את המיטוכונדריה — SS31 ו-MOTS-c בונים אותן. יחד, הם תוקפים את שורש ההזדקנות."
ברפואה וטרינרית מתקדמת, אנו משלבים NAD+ עם פפטידים מיטוכונדריאליים לתוצאות מקסימליות. קרדוסו (2025) מציין כי "NAD+ משתלב בצורה יפה עם MOTS-c ו-SS31 לתיקון מיטוכונדריאלי" [1].
- SS31 (Elamipretide): פפטיד זה חודר למיטוכונדריה ונקשר לקרדיוליפין, מולקולה השומרת על מבנה הקריסטה (הקיפולים הפנימיים). הוא מונע דליפת אלקטרונים ומייעל את ייצור ה-ATP.
- MOTS-c: פפטיד המקודד ב-DNA המיטוכונדריאלי. הוא משפר את הרגישות לאינסולין וממריץ את האנזים AMPK, המעודד יצירת מיטוכונדריות חדשות.
חשוב להבחין בין NAD+ ל-NADH בטיפול. קרדוסו מסביר: "NAD+ הוא הצורה הגולמית שנותנת את המכה למיטוכונדריה. NADH מספק אנרגיה קבועה אבל עובד רק אם יש אנרגיה — צריך את שניהם יחד" [1]. בפרקטיקה הקלינית, אנו ממליצים על פרוטוקולים של 3 עד 6 חודשים כדי לראות שינוי מבני ותפקודי אמיתי בבעל החיים.
8 סיכום ומסקנות (CONCLUSION)
"אל תזלזל בבריאותך — חוסר בקו-אנזים פשוט בגוף יכול לעלות בחיוניות המטופל."
ניהול רמות ה-NAD+ מייצג חזית חדשה ברפואה הווטרינרית האינטגרטיבית. כפי שמסכם קרדוסו (2025), "תהליך האנטי-אייג'ינג קשור לאינספור גורמים. המחסור בקו-אנזים פשוט יכול להביא למספר בעיות בריאותיות והשלכות מטבוליות" [1].
הבנת התפקיד של NAD+ כציר מרכזי בביו-אנרגטיקה מאפשרת לווטרינר לעבור מטיפול בסימפטומים לטיפול בשורש הבעיה. המעבר לתזונה טבעית, העשירה במבשרים וקופקטורים, הוא הצעד היסודי. שילוב של תזונה זו עם תמיכה ממוקדת במיטוכונדריה יכול להחזיר לבעלי חיים קשישים או חולים את החיוניות שאבדה להם.
לסיכום, ה-NAD+ הוא הרבה יותר מתוסף; הוא מדד לבריאות תאית. על הווטרינר המודרני לאמץ אסטרטגיות אלו כדי להבטיח לא רק אריכות ימים, אלא איכות חיים אמיתית עבור המטופלים על ארבע.
רשימת מקורות (REFERENCES)
1. CARDOSO, D. NAD+: the coenzyme that fixes (almost) everything. [S.l.]: Mundo Hormonal, 2025. Available at: https://youtu.be/t1NlydtSodc. Accessed: May 16, 2026.
2. HARDEN, A.; YOUNG, W. J. The alcoholic ferment of yeast-juice. Proceedings of the Royal Society of London. Series B, v. 78, n. 526, p. 369-375, 1906.
3. IMLAI, S. The NAD World 2.0. npj Systems Biology and Applications, v. 2, p. 16018, 2016.
4. YOSHINO, J.; BAUR, J. A.; IMLAI, S. NAD+ intermediates. Cell Metabolism, v. 27, n. 3, p. 513-528, 2018.
5. CANTO, C.; AUWERX, J. NAD+ as a signaling molecule. Current Opinion in Cell Biology, v. 24, n. 2, p. 258-265, 2012.
6. VERDIN, E. NAD+ in aging, metabolism, and neurodegeneration. Science, v. 350, n. 6265, p. 1208-1213, 2015.
7. COVARRUBIAS, A. J. et al. NAD+ metabolism in ageing. Nature Reviews Molecular Cell Biology, v. 22, n. 2, p. 119-141, 2021.
8. KATAYOSHI, C. et al. NMN supplementation improves muscle function in aged mice. Nature Communications, v. 7, p. 13164, 2016.
9. MITSCH, A. et al. NAD+ metabolism and energy homeostasis. Cell Metabolism, v. 22, n. 1, p. 31-53, 2015.
10. NIKIFOROV, A. et al. Pathways of NAD biosynthesis in human cells. Journal of Biological Chemistry, v. 289, n. 34, p. 23713-23725, 2014.
Ver essa foto no Instagram